
We published a paper in February 2018 on the lipid degrading enzyme SAC1. This enzyme takes the phosphate off PI4P lipids, converting them into the less biologically active PI. The enzyme hangs out in the endoplasmic reticulum (ER), is essential for life at the single cell level – yet its substrate is present in other membranes. So what the heck does it do?
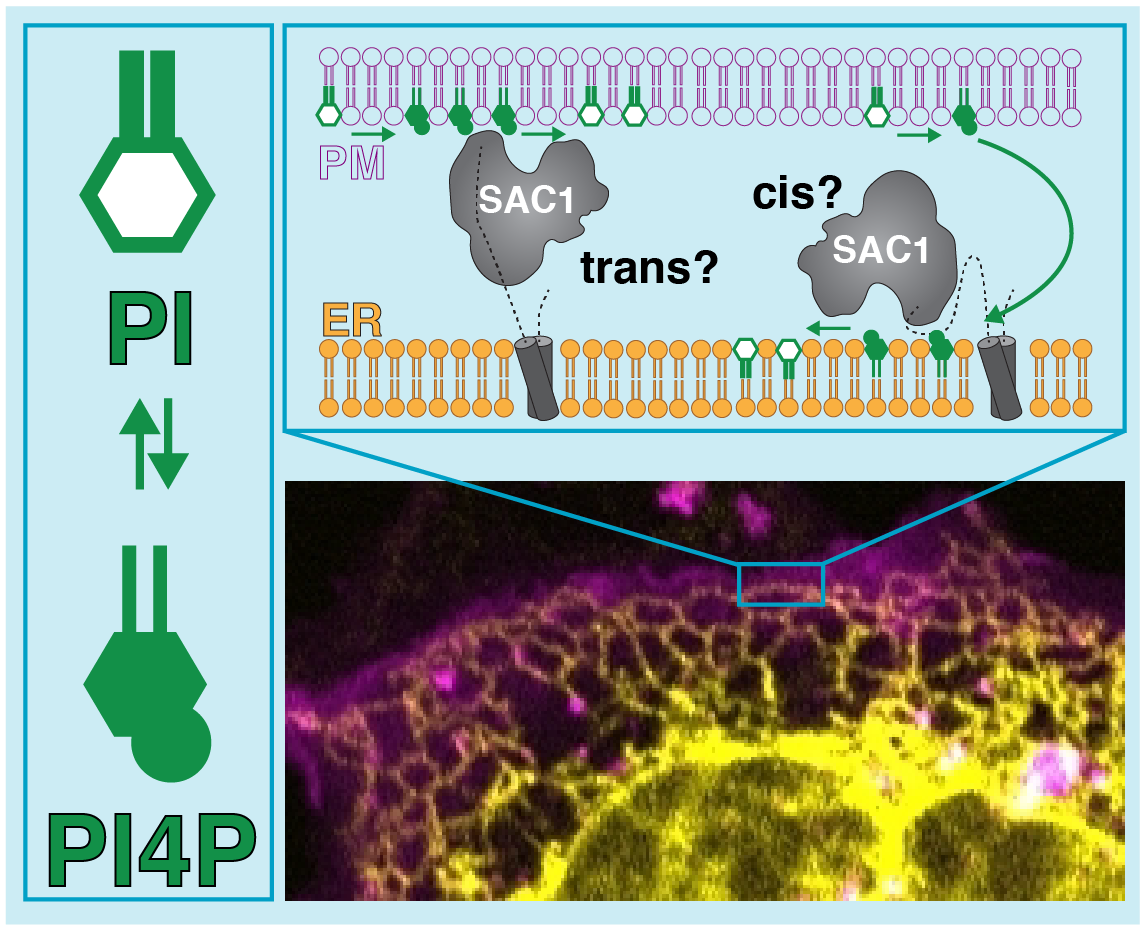
Well, as the picture shows, two possibilities have been thrown out there in the literature. In the first, SAC1 reaches across the gap between ER and a PI4P-containing membrane like the plasma membrane (PM). In the other, PI4P is carried from the PM to the ER so SAC1 can degrade it. We can think of these as “trans” or “cis” modes of action.
So which way does SAC1 actually work? We figured if we block SAC1 activity with a chemical inhibitor like hydrogen peroxide, we could tell the difference: the trans model predicts PI4P would build up in the PM; the cis model predicts PI4P would pile up in the ER (as seen with a fluorescent lipid biosensor). When we did the experiment, this accumulation in the ER was exactly what we saw:

On top of that, the trans model demands that SAC1 works at regions of the ER where it practically touches the PM, the so called membrane contact sites (MCS). We can mark MCS with a fluorescent protein tag – whereas fluorescent SAC1 hangs out all over the ER:

Hmm, so it doesn’t look good for SAC1 to be reaching across the gap and acting in trans mode. As a final test, we asked whether the enzyme was even able to reach across the gap when we forced it to membrane contact sites. It couldn’t, but we could make it: we just had to add a helix to the protein to make the distance between the ER anchor and the catalytic domain longer:
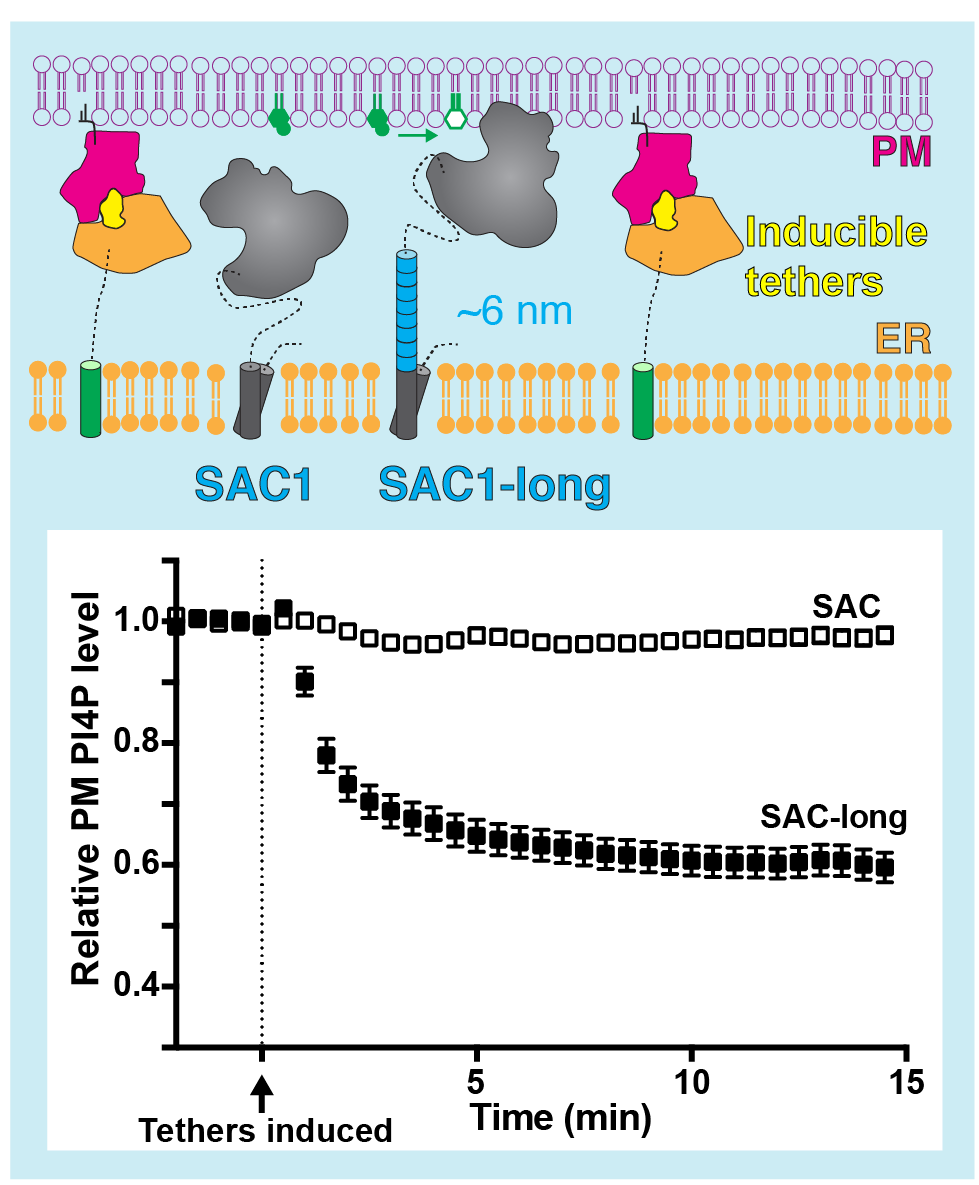
As you can see, this “long SAC1” could now polish off the PM PI4P, whereas the native enzyme could not.
So, native SAC1 seems to be restricted to working in the ER, and having its substrate presented to it by transfer from other membranes. That’s top science-ing Hammond lab. But why did anyone need to know this?
Well, its all about the family of proteins – the ORP proteins – that do the substrate presentation. These can move PI4P as well as other lipids between ER and other membranes like the PM at MCS:

So, now we can see SAC1 has a very important job to do: It breaks down PI4P that is delivered to the ER. Subsequently, the ORPs have to pick up another lipid when they go the other way. Crucially – this other lipid may now be moving against its concentration gradient. This is possible because of the energy released by PI4P flowing down a concentration gradient. The gradient is maintained by SAC1 keeping the PI4P levels low in the ER. You can think of this as being a bit like a water wheel:

What’s next? We’re continuing to explore how these gradients of lipids drive cellular processes, in health and disease. Notably, many single stranded RNA viruses that cause illnesses like the common cold actually seem to hijack this “water wheel”. How exactly they manage this is still not fully understood, but stopping them would have obvious benefits!
You can read the whole paper for free on eLife’s website: https://elifesciences.org/articles/35588
